Fruit Herbivory Alters Plant Electrome: Evidence for Fruit-Shoot Long-Distance Electrical Signaling in Tomato Plants
 Gabriela Niemeyer Reissig1*
Gabriela Niemeyer Reissig1*  Thiago Francisco de Carvalho Oliveira1
Thiago Francisco de Carvalho Oliveira1  Ricardo Padilha de Oliveira1
Ricardo Padilha de Oliveira1  Douglas Antônio Posso2
Douglas Antônio Posso2  André Geremia Parise1
André Geremia Parise1  Dori Edson Nava3
Dori Edson Nava3  Gustavo Maia Souza1
Gustavo Maia Souza1- 1Laboratory of Plant Cognition and Electrophysiology, Department of Botany, Institute of Biology, Federal University of Pelotas, Pelotas, Brazil
- 2Laboratory of Plant Biochemistry and Molecular Biology, Center for Chemical, Pharmaceutical and Food Sciences, Federal University of Pelotas, Pelotas, Brazil
- 3Laboratory of Entomology, Brazilian Agricultural Research Corporation, Embrapa Clima Temperado, Pelotas, Brazil
The electrical activity of tomato plants subjected to fruit herbivory was investigated. The study aimed to test the hypothesis that tomato fruits transmit long-distance electrical signals to the shoot when subjected to herbivory. For such, time series classification by machine learning techniques and analyses related to the oxidative response were employed. Tomato plants (cv. “Micro-Tom”) were placed into a Faraday's cage and an electrode pair was inserted in the fruit's peduncle. Helicoverpa armigera caterpillars were placed on the fruit (either green and ripe) for 24 h. The time series were recorded before and after the fruit's exposure of the caterpillars. The plant material for chemical analyses was collected 24 and 48 h after the end of the acquisition of electrophysiological data. The time series were analyzed by the following techniques: Fast Fourier Transform (FFT), Wavelet Transform, Power Spectral Density (PSD), and Approximate Entropy. The following features from FFT, PSD, and Wavelet Transform were used for PCA (Principal Component Analysis): average, maximum and minimum value, variance, skewness, and kurtosis. Additionally, these features were used in Machine Learning (ML) analyses for looking for classifiable patterns between tomato plants before and after fruit herbivory. Also, we compared the electrome before and after herbivory in the green and ripe fruits. To evaluate an oxidative response in different organs, hydrogen peroxide, superoxide anion, catalase, ascorbate peroxidase, guaiacol peroxidase, and superoxide dismutase activity were evaluated in fruit and leaves. The results show with 90% of accuracy that the electrome registered in the fruit's peduncle before herbivory is different from the electrome during predation on the fruits. Interestingly, there was also a sharp difference in the electrome of the green and ripe fruits' peduncles before, but not during, the herbivory, which demonstrates that the signals generated by the herbivory stand over the others. Biochemical analysis showed that herbivory in the fruit triggered an oxidative response in other parts of the plant. Here, we demonstrate that the fruit perceives biotic stimuli and transmits electrical signals to the shoot of tomato plants. This study raises new possibilities for studies involving electrical signals in signaling and systemic response, as well as for the applicability of ML to classify electrophysiological data and its use in early diagnosis.
Introduction
Plants are sessile organisms with enormous evolutionary success (Bar-On et al., 2018). To cope with unfavorable environmental situations without moving around, they have evolved a body with modular structures and limited tissue differentiation comparing with most animals (Trewavas, 2002; Souza et al., 2018). Thus, since each plant module is able to sense local environmental changes, it is relevant that the many signals coming from the modules be integrated and interpreted effectively, enabling an efficient response in order to maintain the plant surviving as a whole. This is possible through a complex network of short and long-distance signal transduction by hydraulic, chemical, and electrical signaling (Choi et al., 2016).
Electrical signals are not exclusive to organisms that possess nervous systems. In plants, they are derived from changes in cell membrane potentials that can occur due to external tissue stimuli, leading to modifications in ion channels and plasma membrane transporters, which in turn alter membrane voltage by causing an ionic imbalance (Cuin et al., 2018). Studies related to plant electrophysiology generally focus on individual signals of one or few cells, often action potentials (APs) and variation potentials (VPs). However, the analysis of specific signals can underestimate the complexity of many overlapping electrical signals operating simultaneously, which creates a web of systemic information where multiple electrical signals are layered in time and space (De Loof, 2016; Souza et al., 2017). In this sense, it was proposed the term “plant electrome,” which is based on the general definition of “electrome” by De Loof (2016) as the totality of ionic currents of any living entity, from the cell up to the whole organism level. Thus, plant electrome correspond to the plants' bioelectrical activity measured as micro-Voltage changes in stimulated or non-stimulated tissues (Saraiva et al., 2017; Toledo et al., 2019). Recently, plant electrome analysis has been shown as an efficient tool allowing diagnose of different plant stresses (both abiotic and biotic) since the electrome dynamic is sensitive to a plethora of stimuli, exhibiting specific pattern responses recognizable by standard time series analyzes techniques and, specially, by machine learning methods for data classification (Pereira et al., 2018; Simmi et al., 2020; Parise et al., 2021).
The electrical signal seems to be an excellent alternative for fruit signaling to other plant modules and organs due to its inherent capacity for rapid propagation and independence of vascular water mass flow (Fromm et al., 2013; Huber and Bauerle, 2016; Vodeneev et al., 2016). Choi et al. (2017) suggested a general and stress-specific systemic signaling model involving a trio of messengers (ROS, Ca2+, and electrical signals) in which the local stress stimulus triggers changes in membrane potential, increased cytosolic calcium concentration, and activation of RBOHDs (respiratory burst oxidase homolog protein D) mediating the oxidative burst that leads to the accumulation of reactive oxygen species. The association between these signals would generate a wave that quickly spreads throughout the plant in a self-propagating way, through the apoplast or plasmodesmata. Long-distance signaling can result in a systemic response, occurring between tissues of organs with the same function or not. The systemic response of long-distance signaling occurs when an environmental stimulus is perceived in a part of the plant, processed, and the information generated is exported, influencing the behavior of a distant tissue (Choi et al., 2016).
Biotic environmental stressors, such as insect herbivory, promote mechanical damage and facilitate the infection by microorganisms. This type of injury triggers a systemic signaling process, involving hydraulic, chemical, and electrical signals, which in turn can lead, for example, to hormone and oxidative responses (Bi and Felton, 1995; Farmer et al., 2020). The moth Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) is an insect known for its damages to crop production. Its caterpillar present high polyphagy, being able to feed on more than 180 species of plants from different families, including tomato (Solanum lycopersicum L.) (Pratissoli et al., 2015). Also, this species has high fecundity, fast generation time, good ability to disperse in field conditions during the adult stage, and high survivability in adverse environmental conditions (Fitt, 1989; Arnemann et al., 2019). It is widely distributed geographically, being present throughout the Old World and Oceania. Recently, this highly destructive species has reached the Americas, where it was firstly recorded in Brazil and Paraguay in 2013, and in Argentina in 2014 (Czepak et al., 2013; Murúa et al., 2014; Kritikos et al., 2015).
Tomato fruit is a part of the plant that can be damaged by H. armigera caterpillar. Despite of tomato fruits present vascular bundles connecting with the other parts of the plant and the phloem is proposed as the main electrical signaling pathway, information on the role of the electrical signal generated by biotic stimuli perceived in the fruit and its signaling to other parts of the plant is scarce. Thus, since there is a possible route of transmission of electrical signals from the fruit to the rest of the plant, and the injury to the fruit may be signaled to more distant tissues via electrical signaling, it is likely that tomato fruits transmit long-distance electrical signals to the shoot when subjected to herbivory. Studies regarding electrical long-distance signaling under fruit herbivory could be useful to demonstrate that the fruit, as a living and integral part of the plant, is also capable of sending signals when under unfavorable environmental situations, providing to the rest of the plant information allowing to prepare for a possible or imminent environmental adversity.
In this vein, we have tested such possibility analyzing likely changes on plant electrome induced by herbivory in tissues at distance. Moreover, in order to verify long-distance signaling physiological effects, oxidative responses in tissues other than the predated fruits were evaluated.
Materials and Methods
Plant Material and Experimental Conditions
Seeds of tomato cv. “Micro-Tom” wild type (WT) were used. The seeds were germinated in a polystyrene honeycomb germination box, filled with commercial organic substrate, and kept in a germinating chamber (25°C, photophase of 12 h) for 10 days. After this period, the seedlings were transplanted into 0.5 L plastic pots filled with commercial organic substrate and grown under greenhouse conditions. The plants were irrigated on alternate days with 50 mL of tap water. Additionally, 50 mL of nutrient solution (Hoagland and Arnon, 1938) was applied three times a week. The average temperature in the greenhouse during the experimental period was 28.5 ± 12.9°C and the irradiance, from natural light, was 800 μmol photons m−2 s−1.
To carry out the experiment, tomato plants with fruits in two different ripening stages were used: red and mature green. Five tomato plants for each essay were transferred to the laboratory 2 days before the beginning of the experiment to acclimate at room temperature of 25.0 ± 2.0°C and irradiance of 450 μmol photons m−2 s−1 provided by LEDs with full-sunlight spectrum. The caterpillars used in the experiment were in the third and fourth instar. The insects were kept in the laboratory under suitable conditions of temperature (25 ± 2°C), relative humidity (70 ± 15%) and photophase (14 h). The rearing was carried out on an artificial diet according to the methodology developed by Vilela et al. (2014).
The fruits were subjected to herbivory by H. armigera caterpillars, as follows. First the fruits were involved in low-density polyethylene (LDPE) plastic bag, with needle-drilled holes to allow gas exchange. After 2 h acquisition of non-stimulated electrical signals (as described below), a cut in the plastic bag was made for placing the caterpillars on the fruit's surface. Then, the cut was sealed with adhesive tape and the electrical signal acquisition started again for 24 h to gather bioelectrical activity under herbivory. A total of 6 assays were carried out with fruits in the red stage and 4 essays were ran with fruits in the mature green. Each essay was performed with five plants simultaneously. The general experimental setup is showed in Figure 1.
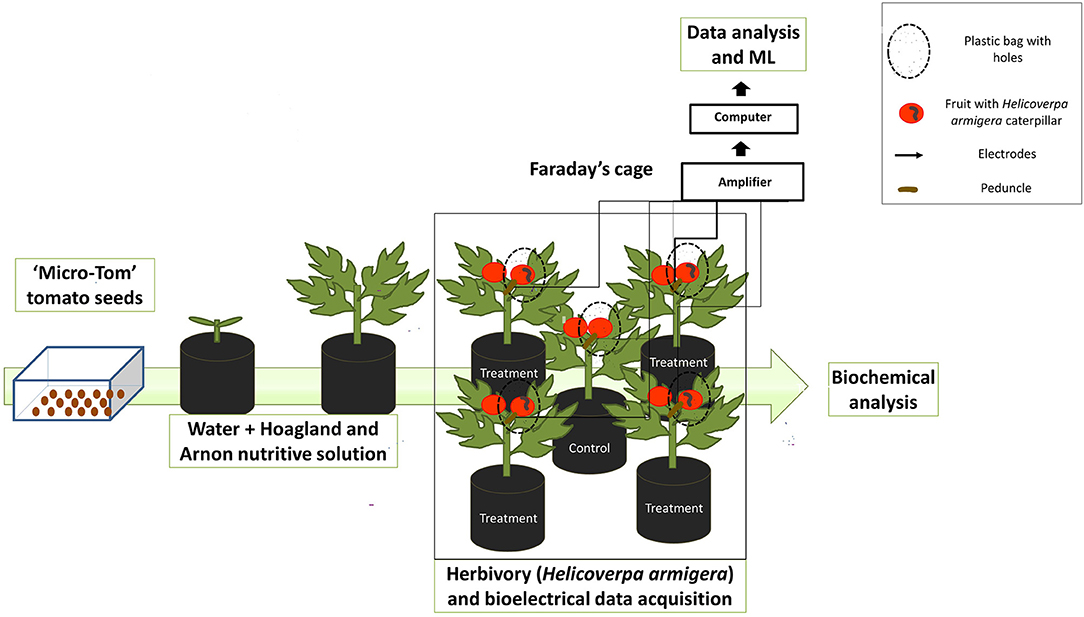
Figure 1. Schematic representation of tomato plant cultivation and experimental set up. Tomato plants were grown until the reproductive stage. Plants with mature green and red fruit were used in the experiment (here we only represented the red fruit). A total of 5 plants were used by assay, 1 control and 4 plants inserted with H. armigera in the fruit. The control was used to collect the fruit and leaves for biochemical analysis and the plants under herbivory were used to the electrophysiological analysis.
Electrophysiological and Biochemical Measurements
Electrome Data Acquisition
Electrical signals were recorded inside a Faraday's cage with the electronic system for data acquisition MP36 (Biopac Systems, Goleta, CA, EUA), composed of four channels with high input impedance (10 GΩ). The signals were acquired by fixing a sampling rate of fs = 62.5 Hz with two filters, one high-pass (0.5 Hz cut-off frequency) and the other low-pass (1.5 kHz cut-off frequency). The bioelectrical runs were analyzed as micro-voltage variation (μV) time series ΔV = {ΔV1, ΔV2, …, ΔVN} in which ΔVi is the difference of potential between the inserted electrodes, scored in each 1/fs time interval, and N is the total length of the series.
In each essay, a pair of needle electrodes (EL452 model; Biopac Systems, Goleta, CA, EUA) was inserted in the peduncle next to the fruit under herbivory (Figure 2) 1 day before of the signal recording for acclimation. The bioelectrical activity was recorded during 2 h before beginning of herbivory and for 24 h after the caterpillars have been placed on the fruits. Additionally, measures with open electrodes (electrodes not attached to plants) were made for the same amount of time that the experiment was carried out to monitoring the equipment noise. For the open electrodes, the signals remained stable throughout the whole period of measurements, showing that the variation in signal complexity was not due to the equipment. Open electrode signals show a typical Gaussian noise with a lower amplitude than the plant signal baseline (Saraiva et al., 2017).

Figure 2. “Micro-Tom” tomato fruit under herbivory. An electrode pair was placed in the peduncle, near the tomato fruit with Helicoverpa armigera caterpillar. Five tomato plants (1 control + 4 herbivory) were kept in Faraday's cage for 24 h, until the end of electrical signal acquisition.
Biochemical Evaluations
Soon after the end of the 24 h electrophysiological measures, red fruits and leaves were harvested from the plants that underwent herbivory, as well as control plants not subjected to herbivory. The samples were immediately stored at −86°C for further biochemical analysis. Prior to the analysis, tomato fruits were macerated in a ball mill with liquid nitrogen and stored in 50 mL falcon tubes in an ultrafreezer (−86°C).
Antioxidant Enzymes
Fruit and leaf samples (± 0.2 g) were homogenized with 100 μM EDTA, 10 mM ascorbic acid, 50% (w:w) polyvinylpyrolidone (PvP) 100 mM, and potassium phosphate buffer pH 6.0 and 7.8 (fruit and leave, respectively). The homogenate was centrifuged at 13,000 × g for 20 min at 4°C and the supernatant obtained was used as a crude enzyme extract (Azevedo et al., 2006). From the same extract, the total soluble proteins content (Bradford, 1976) was determined for expression of enzymatic activity, using a bovine serum albumin standard curve. Mature green fruit and 48 h red fruit were not evaluated due to the difficulty of obtaining the plant material necessary for biochemical analysis from a single “Micro-Tom” tomato plant. The 24 h sample of the red fruit was sufficient for the purpose of this work.
Catalase (CAT—EC 1.11.1.6) activity was determined by incubating fruit and leaf samples with 100 mM phosphate buffer (pH 7.0) and 12.5 mM H2O2 (Azevedo et al., 1998). The enzyme activity was estimated from measurements of H2O2 degradation at 240 nm using ε equal to 36 M−1 cm−1. The results were expressed as μmol H2O2 mg−1 protein min−1 of fresh weight (FW).
Ascorbate peroxidase (APX—EC 1.11.1.11) activity was analyzed using a medium comprised of 37.5 mM phosphate buffer (pH 7.0), 0.25 mM ascorbic acid, and 5 mM H2O2 (Nakano and Asada, 1981). The activity was monitored using ascorbate oxidation rate at 290 nm (ε = 2.80 × 103 M−1 cm−1). The results were expressed in μmol AsA mg−1 protein min−1 FW.
Guaiacol peroxidase (GPOD−1.11.1.7) activity was assayed as described by Azevedo et al. (2006), monitoring the production rate of tetraguaiacol at 470 nm (ε = 26.6 × 103 M−1 cm−1). The reaction medium consisted of 100 mM potassium phosphate buffer (pH 7.0), 0.1 μM EDTA, 5 mM guaiacol, 15 mM hydrogen peroxide, and fruit and leaf extracts. The results were expressed as μmol H2O2 mg−1 protein min−1 FW.
Superoxide dismutase (SOD—EC 1.15.1.1) activity was analyzed using a medium containing 50 mM potassium phosphate buffer (pH 7.8), 14 mM methionine, 0.1 μM EDTA, 75 μM NBT, and 2 μM riboflavin. The enzyme activity was measured at 575 nm, and 1 unit of activity was defined as the amount of enzyme required to inhibit the reduction of nitroblue tetrazolium by 50 % (Giannopolitis and Ries, 1977).
Hydrogen Peroxide and Superoxide Anion
The determination of hydrogen peroxide was performed using a methodology described by Velikova et al. (2000). Tomato fruit (± 0.2 g), previously macerated and cold stored, and leaf (± 0.2 g) were homogenized in 0.1% trichloroacetic acid (TCA). The homogenate was centrifuged at 13,000 × g at 4°C for 20 min and the supernatant obtained was added to the reaction medium, which was composed of 10 mM potassium phosphate buffer (pH 7.0) and 1 M potassium iodide. The absorbance was read at 390 nm. The content was determined by a standard curve prepared with known concentrations of H2O2 and the results expressed in μmol H2O2 g−1 FW.
Superoxide anion content was determined using the methodology of Elstner and Heupel (1976). Approximately 0.2 g of tomato fruit and leaves were homogenized in a 65 mM phosphate buffer (pH 7.8) and centrifuged at 5,000 × g at 4 °C for 10 min. The supernatant obtained was mixed with 65 mM phosphate buffer (pH 7.8) and 10 mM hydroxylamine. Then the reaction medium was immediately placed in a water bath at 25°C for 20 min. Thereafter, 17 mM sulfanilamide and 7 mM α-naphthylamine were added, and again the reaction medium was incubated at 25 °C for 20 min. The absorbance of the solution was measured at 530 nm. A standard curve of sodium nitrite (NaNO2) was used to calculate the superoxide anion content. The results were expressed in μmol g −1 FW.
Data Analysis
Statistical Analysis of Biochemical Measurements
Five replicates (n = 5) of red fruit (control and 24 h under herbivory) and leaves (control and 24 and 48 h under herbivory) were analyzed by one-way analysis of variance (ANOVA). When F was significant, the treatment means were compared with Student's t-test (p ≤ 0.05). Statistical analyses and graphics were performed using Sigmaplot 12.0 (Systat Software Inc., USA).
Machine Learning Classification Methods on Electrome Time Series
The characterization of the electrophysiological time series collected before and after herbivory was carried out. We obtained the following time series: 16 series of 2 h for green fruit plants before herbivory; 16 series of 24 h for green fruit plants after herbivory; 22 series of 2 h for red fruit plants before herbivory; 22 series of 24 h for red fruit plants after herbivory. These series were filtered and divided in order to increase sample space, which resulted in 150 samples for green fruits before herbivory (GB), 150 samples for green fruits after herbivory (GA), 220 samples for red fruits before herbivory (RB) and 220 samples for red fruits after herbivory (RA). The samples are subdivisions of the series, in shorter time series, in order to increase the sampling space. All the time series were divided into 10 interchangeable parts between them with a lag of 30%, meaning that each part overlaps the other. In the end, each sample had ~45,000 (2 h) and 540.000 points (24 h). The time series were filtered through notch filter (notch = 60) and band pass_filter (low = 32 and high = 0.5) with the function applies filtfilt from the scipy.signal library.
For the generation of the features to feed machine learning (ML) algorithms, the Fast Fourier Transform (FFT), Spectral Power Density (PSD), Wavelets, and Approximate Entropy (ApEn) were calculated for each of the samples (Saraiva et al., 2017; Parise et al., 2021). For FFT, PSD, and Wavelets we calculate the average, the maximum value, the minimum value, the variance, skewness, and kurtosis. Moreover, in order to decrease the computational cost speeding the analysis, we decided to reduce these features by calculating a Principal Component Analysis (PCA) in the descriptive analysis calculated for FFT, PSD, and Wavelets (Parise et al., 2021). Thus, we obtained the first three PCA components (PCA1, PCA2, and PCA3). Finally, we obtained four features: ApEn, PCA1, PCA2, and PCA3.
ML Training and Testing
Considering the inherent complexity of electrome data, different methods of ML classification have been used to ensure more reliability to the results of classification (Pereira et al., 2018). Herein, ML training was performed in a supervised manner. All data were calculated using numpy.random.seed with seed = 42. The values for the hyperparameters were obtained from the literature and from a previous evaluation on the data sets. The hyperparameters that obtained the best levels of accuracy were maintained for all the underlying analyzes. When the default hyperparameters reached the best results, they were kept. The different classification processes used are detailed hereafter.
- Nearest Neighbors: (n_neighbors = 2) this method uses the inputs closest to each other to group and classify each group (Cover and Hart, 1967);
- Linear Support Vector Machine (Linear SVM): (max_iter = 100.000 and tol = 1e-1) the Linear SVM method predicts through the linear regression the position of each entry and consequently which group the entry belongs to Fan et al. (2008);
- Radial Basis Function SVM (RBF SVM): (gama=1e-2) it is similar to the Linear SVM method, however, the RBF uses another strategy to plot the hyperplane decision boundary between classes, such as curves in addition to lines (Chang et al., 2010);
- Gaussian Process: (max_iter_predict = 150 and multi_class = one_vs_one) this method uses input interpolation and Gaussian probabilistic statistics to predict results (Rasmussen and Williams, 2006);
- AdaBoost (Adaptive Boosting): (n_estimators = 100) this method uses a meta-heuristic algorithm to adapt the classification of each entry (Freund and Schapire, 1997);
- Naïve Bayes: (var_smoothing = 1e-7) uses Bayes' theorem to classify each entry (Russell and Norvig, 2003);
- Quadratic discriminant analysis (QDA): (default) assumes that the inputs are normally distributed, however, there is no need to assume that the covariance of each group is identical (Kim et al., 2011);
- Decision Tree Classifier: (max_depth = 10 and min_samples_leaf = 64) creates a decision tree based on the relevance of each feature so it classifies the entries using the paths previously created (Breiman et al., 1984);
- Neural Net: (default) This method uses a network of nodes specialized in classifying specific features giving weights that at the end are added up and return a classificatory result for each group. The model used in this work was the MLPClassifier, which optimizes the log-loss function using LBFGS (Windeatt, 2006).
Additionally, the DummyClassifier method was used as a control, since it classifies each group without an intelligent strategy, being guided by chance. Intelligent methods that have their accuracy values (considering their respective deviations) close to that found by the Dummy were considered not capable of classifying the inputs.
The Stratified KFold (n_splits = 4 and shuffle = True) and Cross Validate methods (Forman and Scholz, 2010; Adagbasa et al., 2019) were used to minimize the effect of chance on the distribution of data used for training and testing.
The code libraries used were: Numpy and Pandas for data manipulation; Scipy, Obspy and Math for mathematical calculations; Matplotlib for creating graphics; Sklearn and Statsmodels for machine learning. To test our hypothesis, the groups GB × GA, RB × RA, RB × GB, and RA × GA were classified using the methods described above. The methodology used here has been previously described by Parise et al. (2021). All the algorithms were running in Python.
Results
Electrome Classification by ML Before and After Fruit Herbivory
Figure 3 shows the decision making of each method to classify the GB and GA groups. The method that obtained the best accuracy value was the Gaussian Process with 98.21% ± 4.64 of accuracy (Figure 3). The Dummy method found a value of 66.67% ± 0.88. We see clearly that ML was able to classify the electrome among the groups. Several models have achieved high accuracy values (Figure 3), demonstrating that the GB and GA groups are easily classified. These data indicate that the herbivore in the mature green fruit considerably modifies the electrome registered in the peduncle, demonstrating that the electrical signal generated in the fruit was transmitted to the rest of the aerial part of the plant.
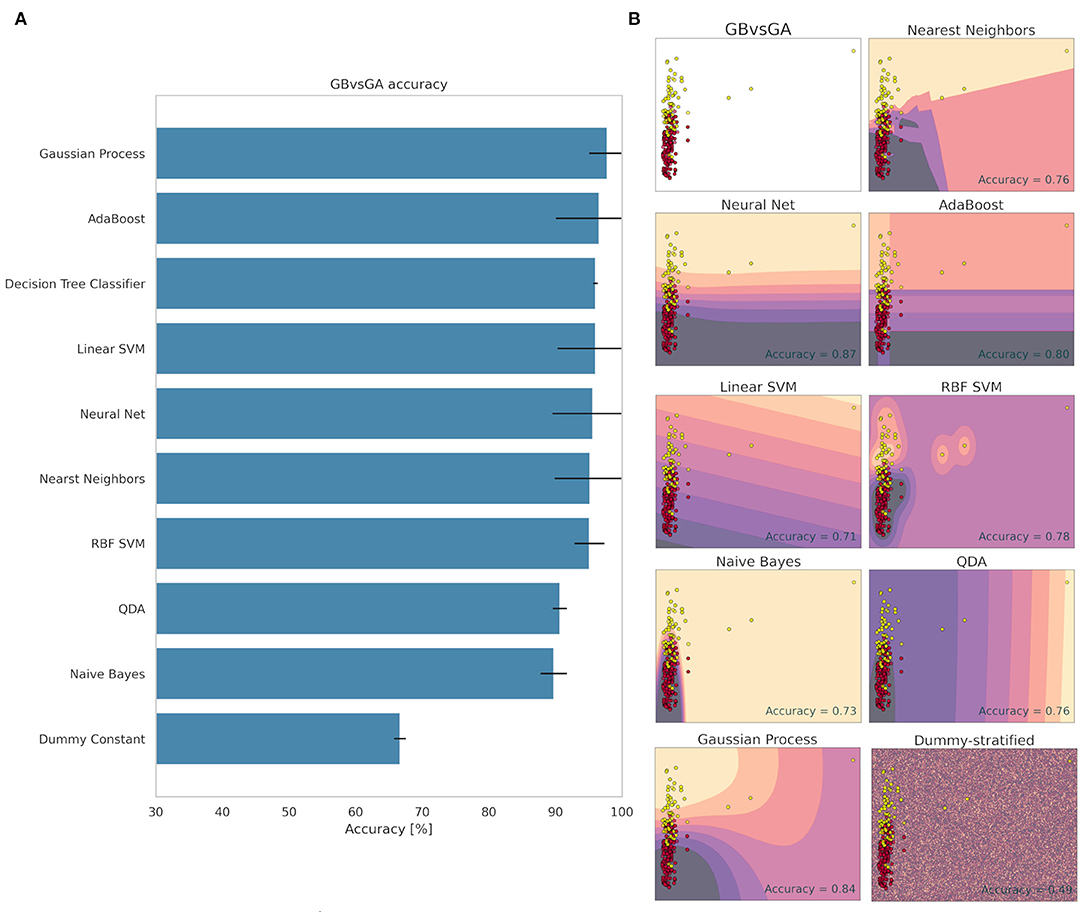
Figure 3. Accuracy (%) and standard deviation (A) of the Machine Learning models used to classify GB and GA groups. Scatter plots (B) showing a sample space plan used in ML training for the GB × GA distribution. Red dots: GB; Light yellow spots: GA. The regions are the demarcations made by ML to indicate levels of certainty of classification. The darker it is, the more certain it belongs to the GB group. The clearer the more certainty of belonging to the GA group. The subtitle of each graph indicates the method used and in the lower right quadrant is the accuracy found.
The decision-making used by the methods to classify between the RB and RA groups is shown in Figure 4. The best method was AdaBoost with 99.72% accuracy (± 1.73). All other methods reached values above 98% accuracy (Figure 4), not including the Dummy, which obtained an accuracy of 66% ± 0.01. This high value demonstrates how easily the RB and RA groups were differentiated, indicating that the red fruit also changes the electrome, emitting electrical signals to the plant when subjected to herbivore on the fruit.
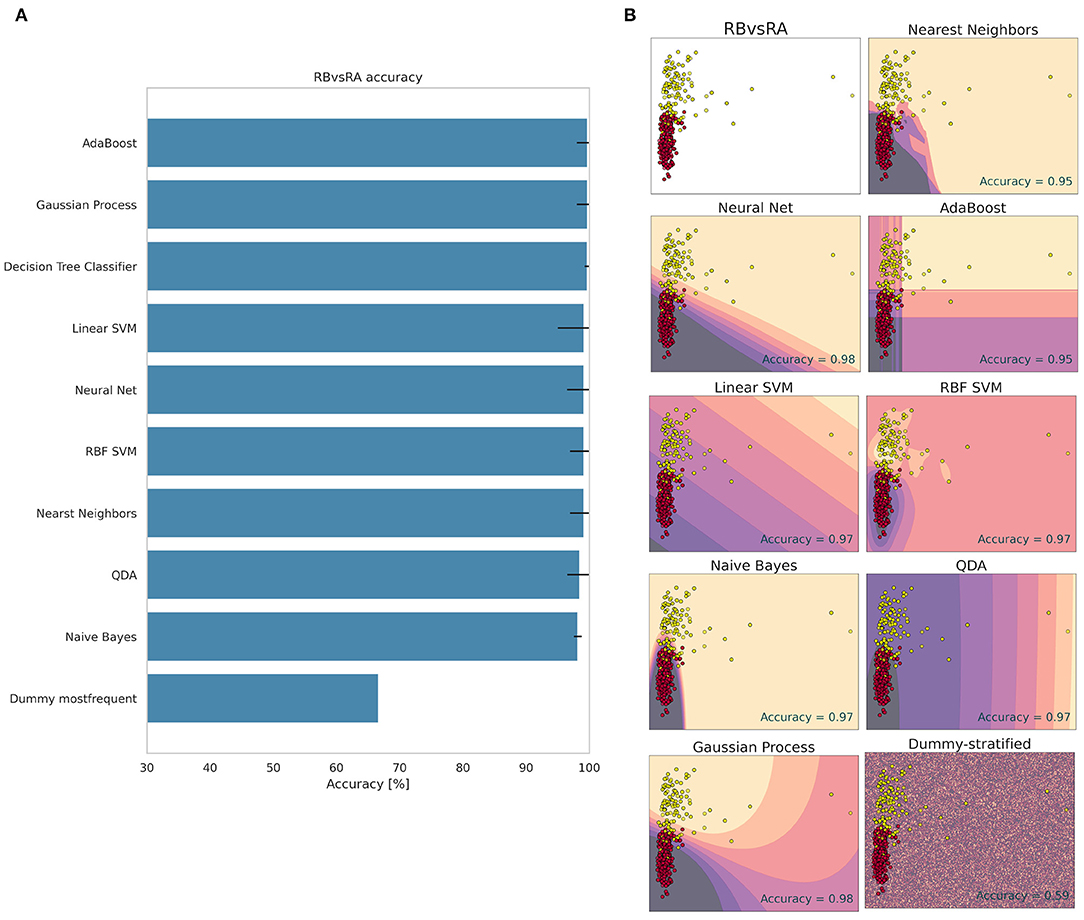
Figure 4. Accuracy (%) and standard deviation (A) of the Machine Learning models used to classify RB and RA groups. Scatter plots (B) showing a sample space plan used in ML training for the RB × RA distribution. Red dots: RB; Light yellow spots: RA. The regions are the demarcations made by ML to indicate levels of certainty of classification. The darker it is, the more certain it belongs to the RB group. The clearer the more certainty of belonging to the RA group. The subtitle of each graph indicates the method used and in the lower right quadrant is the accuracy found.
The decision making for the classification of the RB and GB groups can be seen in Figure 5. The method that obtained the best accuracy for classifying the groups was the Gaussian Process with 90.55% ± 4.39 of accuracy. The Dummy obtained 59.46% accuracy ± 0.01. These values indicate that the ML was able to classify the groups with great accuracy and reliability (Figure 5). The other methods obtained lower accuracy values than the Gaussian Process (Figure 5). This result indicates that there is a difference between the electrome of the mature green and the red fruit, even before subjecting the plant to an external stimulus, and that this difference is reflected in measurements made on the fruit's peduncle. This finding corroborates Reissig's et al. (under review) study, where the electrome of ripening tomato fruits was analyzed.
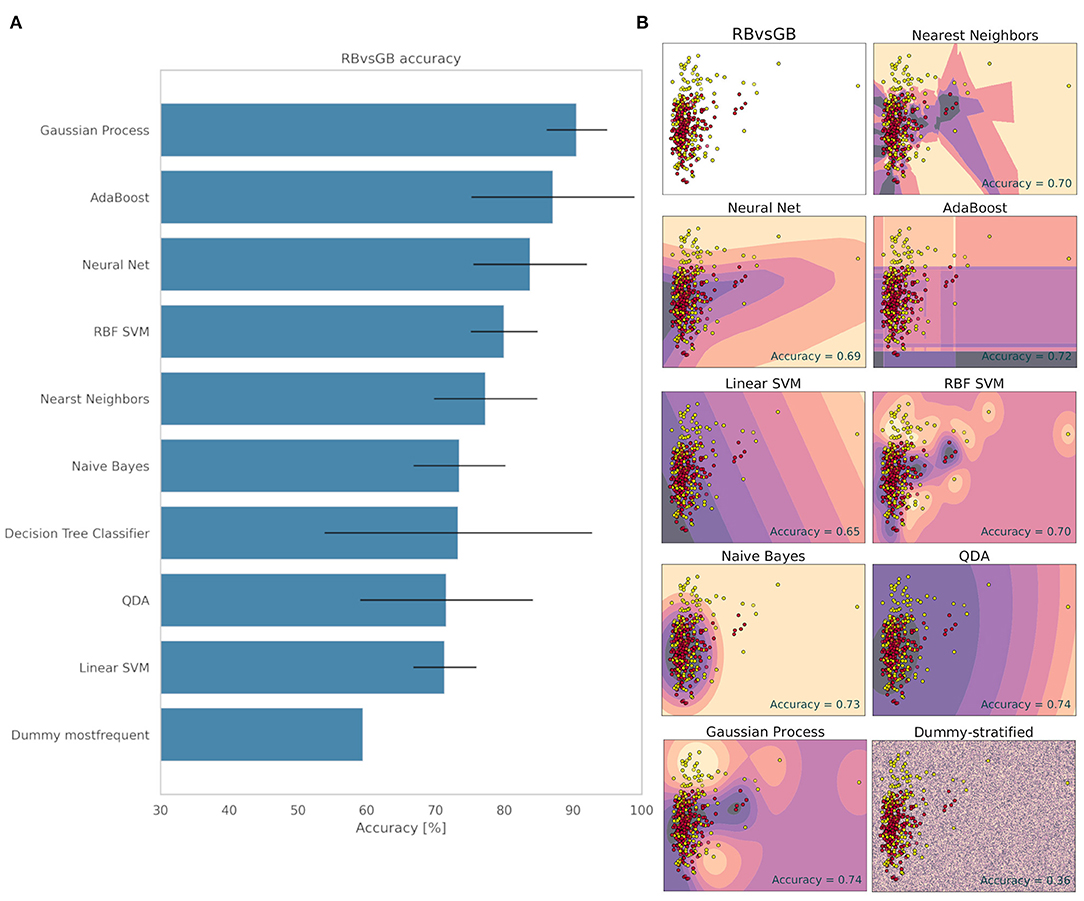
Figure 5. Accuracy (%) and standard deviation (A) of the Machine Learning models used to classify RB and GB groups. Scatter plots (B) showing a sample space plan used in ML training for the RB × GB distribution. Red dots: RB; Light yellow spots: GB. The regions are the demarcations made by ML to indicate levels of certainty of classification. The darker it is, the more certain it belongs to the RB group. The clearer the more certainty of belonging to the GB group. The subtitle of each graph indicates the method used and in the lower right quadrant is the accuracy found.
The strategies used to classify the RA and GA groups are shown in Figure 6. In a visual analysis of the scatter plot, one can observe how difficult is separate it in two groups. The samples have little variation from each other, this indicates a similarity between both groups, which made the ML learning difficult. The method that obtained the highest accuracy for classifying the RA and GA groups was AdaBoost with 70.65%. However, its deviation was 20.74%, which is remarkably high, indicating a significant dependence on data separations and, therefore, low learning reliability. The second-best method was Neural Net (66.38% ± 9.37). Nevertheless, the Dummy method obtained 61.65% accuracy with 5.12% of standard deviation (Figure 6). Considering the standard deviation, it can be said that none of the methods learned how to differentiate the RA and GA groups. This result shows that the electrome after the herbivore is similar for both the red and mature green fruits, standing out from the differentiated signal observed between the mature green and red fruits before herbivory.
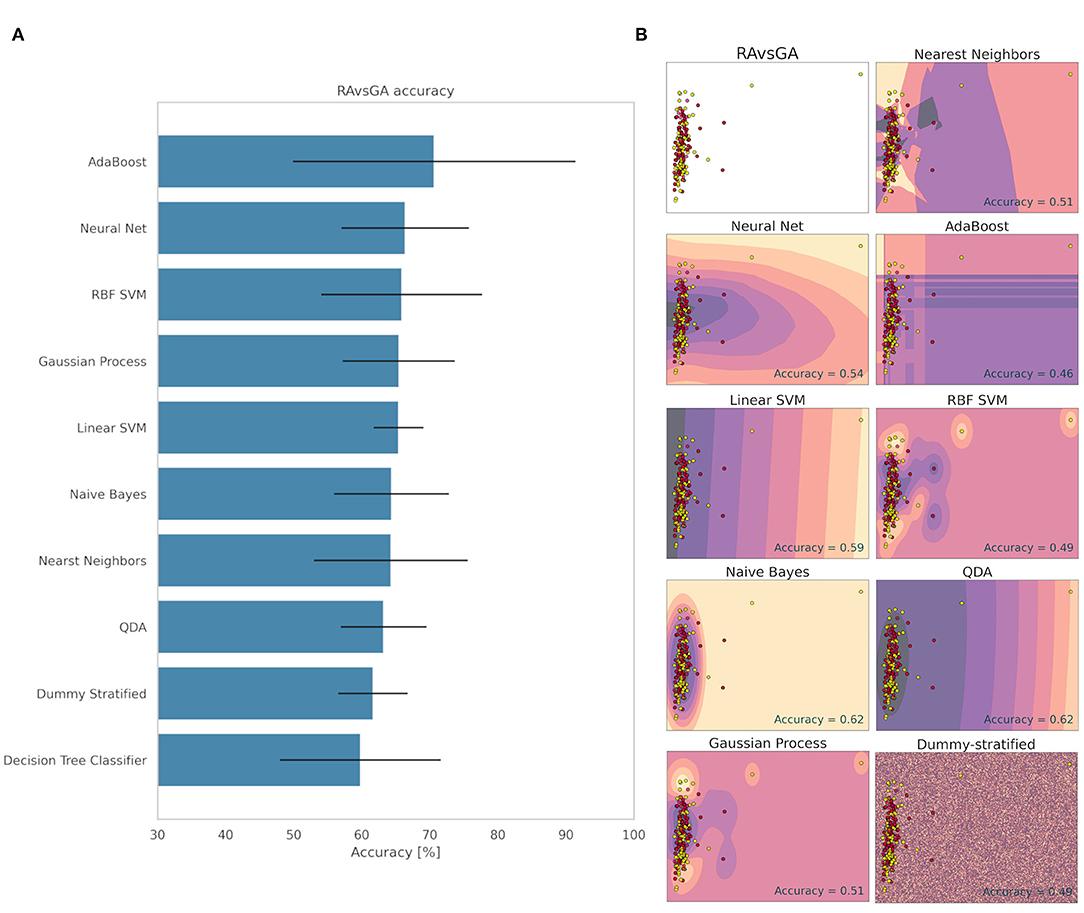
Figure 6. Accuracy (%) and standard deviation (A) of the Machine Learning models used to classify RA and GA groups. Scatter plots (B) showing a sample space plan used in ML training for the RA × GA distribution. Red dots: RA; Light yellow spots: GA. The regions are the demarcations made by ML to indicate levels of certainty of classification. The darker it is, the more certain it belongs to the RA group. The clearer the more certainty of belonging to the GA group. The subtitle of each graph indicates the method used and in the lower right quadrant is the accuracy found.
Herbivory-Induced Systemic Response
To evaluate whether the fruit herbivory generated a systemic oxidative response and signaling in tomato plant, ROS and antioxidant enzymes analyses were carried out (Figures 7, 8). Hydrogen peroxide (H2O2) content was significantly (p ≤ 0.05) higher in the fruit close to the one that was subjected to herbivory (Figure 7). The same was observed in the leaf 24 h. Comparing the leaves 24 and 48 h of the herbivory treatment, it is possible to notice a decrease in the H2O2 concentration in the latter. Regarding superoxide anion content, there was no significant difference (p ≥ 0.05) between control and herbivory in the fruit, nor in the leaf 24 h (Figure 7). Nevertheless, there was a decrease in the superoxide anion content in the leaf 48 h under herbivory, differing significantly (p ≤ 0.05) from the control, which maintained the same level as in the 24 h.
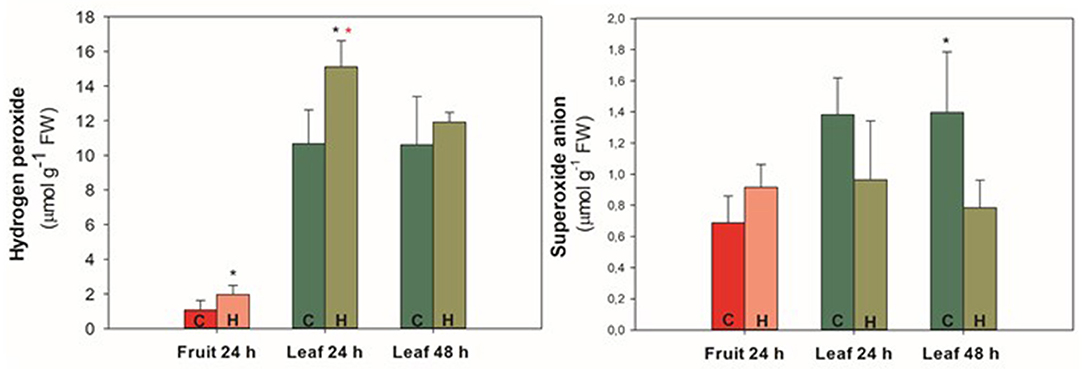
Figure 7. Hydrogen peroxide and superoxide anion content of “Micro-Tom” tomato fruit and leaves during 24/48 h of herbivory treatment (H. armigera). Values represent the mean ± SD (n = 5). Black * represents statistical difference between control and herbivory 24 h or control and herbivory 48 h. Red * represents statistical difference between control 24 and 48 h or herbivory 24 and 48 h. C, control treatment; H, herbivory treatment.
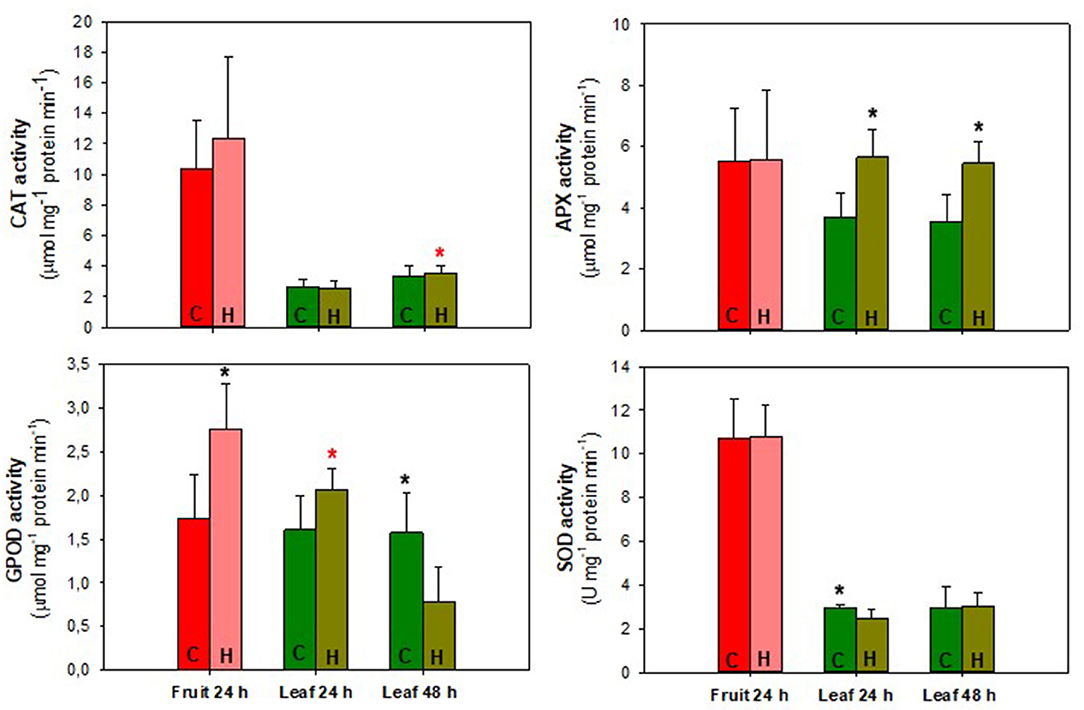
Figure 8. Antioxidant enzymes activity of “Micro-Tom” tomato fruit and leaves during 24/48 h of herbivory treatment (H. armigera). Values represent the mean ± SD (n = 5). Black * represents statistical difference between control and herbivory 24 h or control and herbivory 48 h. Red * represents statistical difference between control 24 and 48 h or herbivory 24 and 48 h. C, control treatment; H, herbivory treatment.
There was no significant difference (p ≥ 0.05) between control and herbivory treatment for fruit and leaves. A significant increase (p ≤ 0.05) in CAT activity was only observed when the leaf 24 h was compared to the leaf 48 h, both under fruit herbivory conditions. Regarding APX activity, there was no significant difference (p ≥ 0.05) in the fruit 24 h between control and herbivory. However, both for 24 and 48 h, APX activity was higher in the leaves belonging to the fruit herbivory treatment. GPOD was the only enzyme that showed higher activity in the fruit 24 h, compared to the control. The leaves of the control treatment maintained practically the same GPOD activity between 24 and 48 h. Leaves belonging to the fruit herbivory treatment presented higher activity in the 24 h. In the 48 h of herbivory, the activity decreased, being significantly lower than the control (p ≤ 0.05).
SOD activity was practically the same between 24 and 48 h in the leaf control treatment. In the 24 h, the control leaf showed an activity significantly (p ≤ 0.05) higher than the leaf of the herbivory treatment. When the analysis was performed on the leaf 48 h, it was observed that the activity increased and there was no difference compared to the control. There was no significant difference (p ≥ 0.05) in SOD activity between fruits. In all analyses performed, there is a stability between the control leaves 24 and 48 h, with no significant difference (p ≥ 0.05) in any of the cases. Regarding herbivory treatments, there is a fluctuation in concentration and activity, effectively demonstrating the influence of herbivory on the fruit in the oxidative response of different organs.
Discussion
Plant herbivory caused by insects is responsible for large production losses worldwide (Arnemann et al., 2019). It is of the utmost importance to know all aspects involving plant-insect interactions, from the ecological to the physiological perspective, in order to develop efficient strategies to deal with damages caused by them (Burkepile and Parker, 2017). Several studies have been carried out to understand the mechanisms behind signaling and systemic responses of plants under herbivory. Due to its inherent sessility, plants possess a plethora of mechanisms and molecules to deal with herbivory, such as RNA silencing, peptides, and specialized metabolites. The complexity of plant herbivory signaling arises from the huge number of plant-insect interactions, resulting in a panoply of defense compounds and systemic signaling elicitors. Exogenous ATP, glutamate, cell wall oligogalacturonides are examples of herbivory systemic response elicitor (Arimura, 2020; Farmer et al., 2020). In addition to the elicitors, a recent study demonstrates that ethylene response factors 15 and 16 trigger jasmonate biosynthesis in tomato during herbivore resistance (Hu et al., 2021). These receptors, as well as ATP, glutamate, and cell wall oligogalacturonides, are also present in fruits and involved in ripening and defense responses (Gao et al., 2020).
In the last decade, many studies have focused on the involvement of the electrical signals in the systemic signaling of plants under herbivory, especially due to its rapid propagation. Also, due to the involvement of calcium and the depolarization of the plasma membrane caused by this sort of injury. The calcium ion (Ca2+) is a universal signaling element in plant cells, as well as in all eukaryotic cells, and is intricately linked to the electrical signal. Recent studies are suggesting the ion channels of GLR (Glutamate Receptor–Like) family and CNGCS (Cyclic Nucleotide Gated Channels) are involved in systemic signaling (Toyota et al., 2018; Breeze, 2019). The H+-ATPase is a proton pump ATPase that couples ATP hydrolysis with the transport of protons out of the cell, generating a proton motive force. It supports the uptake of ions and influences on the cytoplasmatic and apoplastic pH (Morsomme and Boutry, 2000). The electrogenic pumps are assumed to participate in action potentials (APs) generation. H+-ATPases are reversibly inactivated by calcium (Ca2+) influx at the beginning of AP generation, being activated in the repolarization phase (Vodeneev et al., 2016). The AHA1 proton pump is involved in the herbivory-induced electrical signals, especially the slow wave potential (SWP). This proton pump is necessary for SWP repolarization in Arabidopsis (Kumari et al., 2019; Farmer et al., 2020). All the components previously mentioned, which are involved in the electrical signal and systemic response to herbivory, and which are generally studied in Arabidopsis thaliana plants, are also present in fruits (Ewing and Bennett, 1994; Aouini et al., 2012; Zhou et al., 2020), supporting the possibility of fruit-shoot long-distance electrical signaling.
In terms of individual electrical signals, the system potential (SP) (Zimmermann et al., 2016) and the slow wave potential (SWP) (Farmer et al., 2020) are the most reported signals involved in biotic stress. Although studies have already shown the existence of these signals during herbivory, much still need to be done to understand the information that these signals carry and how these signals are transduced. It is not possible to identify specifically which individual electrical signal is pre-dominant in the present study. Nonetheless, our finding showed the transmission of the signal generated in the fruit to the peduncle, and probably to rest of the plant. The ML was efficient to differentiate the signal before and after herbivory (Figures 3, 4), both for mature green and red fruit. This finding opens new opportunities for studies regarding fruit-to-shoot signaling.
Farmer et al. (2020) proposed a model for slow wave potential (SWP) propagation in Arabidopsis under herbivory. In this model, in a nutshell, the wounding caused by caterpillar will promote loss of phloem pressure and a loss of tension in the xylem. Then, due to the pressure changes in the vessels, the elicitors would be carried into and along the xylem, interacting with cells and promoting more depolarization. This first stage would be predominantly non-electrical. After these events, the electrical part of the SWP propagation would begin. The subsequent stage is characterized by an increase in cytosolic calcium and synthesis of jasmonate from precursors produced by LOX6. The jasmonates produced would then be exported to activate defense responses. The authors argue that both xylem and phloem are important to SWP propagation. It was not mentioned in the study the possibility of this model in fruits under herbivory. However, it seems quite reasonable to extend this model to this part of the plant, since this structure is connected via peduncle (pedicel) with the rest of the plant.
Fruits such as tomato are connected to the shoot of the plant through vascular bundles. The xylem and phloem are the structures through which the fruits will receive water, carbohydrates, calcium, among other compounds (Rancic and Pecinar, 2010). In addition, phloem is proposed as the main route of transmission of electrical signals, due to the properties of its cells. In relation to the other cells, the sieve tube elements have great length and sieve plates with large pores, which decrease the electrical and hydraulic resistance between one cell and another. They are also devoid of vacuole, nucleus, and plastids (Hedrich et al., 2016). Although fruits, especially the fleshy ones, are modules of the plant with the purpose of being consumed to disperse the seeds—when already ripe—, they are still capable of perceiving environmental disturbances and generating a signaling and systemic responses in the plant. It is known that even in the post-harvest the fruits are responsive to different stimuli and able to adjust their metabolism (Pedreschi and Lurie, 2015; Reissig et al., 2018).
The oxidative response has already been reported in plants under herbivory and wounding. In a study with unripe wounded avocado fruit, decreased activity of CAT and SOD was observed after 24 h of injury (Castro-Mercado et al., 2009). In our study, there was no significant difference (p ≥ 0.05) between the control and the herbivory treatment for these same enzymes in the fruit and leaf (Figure 8), except in the leaf 24 h, where the SOD activity for herbivory treatment was lower than the control, similar to the observed by Castro-Mercado et al. (2009). It should be noted that the fruits analyzed in our study were not the same as those that suffered herbivory. Interestingly, as in the study with avocado, we observed a considerable increase in fruit GPOD activity, being the only antioxidant enzyme that increased its activity in the fruit belonging to the plant that underwent herbivory. Regarding leaf enzyme activity, APX was the one that stood out, having its activity greater than the control during 24 and 48 h of herbivory (Figure 8). In a study on leaf oxidative stress in soybean plants, induced by Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae) herbivory, an increase in APX enzyme activity was also observed after 3 days of treatment (Bi and Felton, 1995). Duffey and Stout (1996), wrote that as a direct defense to herbivory in tomatoes, antioxidant enzymes have a reduced effect. However, together with alkaloids, proteinase inhibitors and phenolic compounds there is a synergistic effect, affecting the insect during its digestive process and metabolism.
The H2O2 was the ROS that showed the highest accumulation during herbivory, especially in the first 24 h, both the fruit and the leaf (Figure 7). Superoxide anion decreased after 48 h of herbivory in tomato plant leaf and there was no significant difference (p ≥ 0.05) between fruit control and herbivory treatment (Figure 7). Due to its high stability and free diffusion, H2O2 plays an important role in plant defense responses. One of the main producers of H2O2 in plant cells under the most different stresses is the NADPH oxidase (also known as respiratory burst oxidase homolog—RBOH) present in the plasma membrane. It generates superoxide anion, which in turn is converted in H2O2 by SOD activity (War et al., 2012; Liu and He, 2016). We did not evaluate ROS right at the beginning of the herbivory on the fruit. Probably, the concentration of superoxide anion was higher in the beginning, before being converted to H2O2. Perhaps this is the reason why the concentration of superoxide anion was equal or lower than the control. In addition to stress response, ROS are also involved in systemic signaling, acting together with phytohormones and other signaling molecules (Fichman and Mittler, 2020).
In the present study, we analyzed components involved in the oxidative response as a marker, to demonstrate that the stimulus provoked in the fruit by the H. armigera caterpillar was sufficient to generate a response in more distant organs. Here we cannot make a direct link of the electrical signal generated in the fruit with the observed response, but it is likely that it is coupled with other forms of herbivory signaling already established (e.g., jasmonates, salicylates, Ca2+). The fact that herbivory on the fruit has led to a response in different organs shows the importance of studying and understanding the signaling that can be generated from the fruit. Valoy et al. (2019) observed in their studies that leaf herbivory affects the correlation between fruit traits (such as weight, number of seeds, and sucrose), and the magnitude and variation of the damage that influences it. It is reasonable to assume that something similar can occur when another fruit is being preyed, instead of the leaf. Although the fruits do not depend directly on each other as they depend on the leaves to receive photoassimilates, it is possible, as shown here, that biochemical changes involved in systemic responses between them do occur.
Machine learning techniques have emerged as a powerful tool for several studies in the field of plant physiology, both for technical application and as a tool to help test scientific hypotheses (Pereira et al., 2018; Tran et al., 2019; de Medeiros et al., 2020; Simmi et al., 2020). In addition to the central hypothesis of this research, the use of ML allowed us to observe differences in the electrome of tomato plants when the fruits were in different ripening stages. The ML was able to differentiate tomato plants with green fruits from ripe ones before herbivory, presenting a high accuracy (Figure 5). However, in the presence of H. armigera, it was not possible to differentiate between them. The signals generated by herbivory were predominant, making it possible for them to be easily classified by ML. This finding may bring new insights into how electrical signals are involved not only in external disturbances but also in signaling physiological processes. Our research group also carried out experiments focusing on the ripening of climacteric fruits, where a difference was observed in the electrome between the ripening stages of harvested tomato fruit (Reissig et al., under review).
Arimura (2020) highlights the myriad of agricultural applications provided by new insights regarding studies into the cellular pathways by which plants sense elicitors and elicit defense responses against herbivore. Here, we also envision potential applications of electrical signal analysis, especially associated with machine learning techniques. Regarding other sources of biotic stress, a recent study with a pathogenic fungus showed that the infection by Oidium neolycopersici altered the electrophysiological dynamics of the tomato plant. These changes were detected far from the infection site and before any visual symptom, raising the possibility of using electrome analyses for early diagnosis of plant diseases (Simmi et al., 2020). Tran et al. (2019) demonstrated this possibility by studying abiotic environmental stimuli. They proposed the electrophysiological evaluation of tomato plants status using supervised machine learning. It would be achieved through an electrophysiological sensor that allowed the measurement of real-time electrical signals related to the plant water status in the field, without a Faraday cage. This possibility can be extended to biotic stress factors, enabling the development of sensors and ML techniques that detect and manage to differentiate, in the case of herbivory, the insect that is causing injury in a crop production early in the infestation, allowing less aggressive measures and more accurate insect management.
Concluding, the tomato fruit is a plant organ capable of sensing biotic stimuli and transmitting electrical signals to the shoot of tomato plants. Besides, the signaling between fruit and shoot triggered biochemical processes related to biotic stress responses in different leaves and fruits. Finally, we observed that the electrome in the peduncle was different between mature green and red fruits, indicating that electrical signals might be involved in the ripening process. This study raises new possibilities for studies involving electrical signals in signaling and systemic responses, as well as for the applicability of ML to classify electrophysiological data and its use in early diagnosis. It could contribute to a more sustainable crop and food production.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Author Contributions
GR: conceptualization, methodology, formal analysis, investigation, data curation, and writing—original draft preparation. TO: formal analysis, investigation, data curation, and writing—original draft preparation. RO: investigation and Helicoverpa armigera rearing. DP: investigation and data curation. AP: writing—reviewing and editing. DN: Helicoverpa armigera rearing and resources and writing—reviewing and editing. GS: resources, writing—reviewing and editing, and supervision. All authors contributed to the article and approved the submitted version.
Funding
This study was supported in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brasil (CAPES), Finance Code 001. Authors are also grateful to Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the financial support provided (401722/2016-3). GS was CNPq fellow (302715/2018-5).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Adagbasa, E. G., Adelabu, S. A., and Okello, T. W. (2019). Application of deep learning with stratified K-fold for vegetation species discrimation in a protected mountainous region using Sentinel-2 image. Geocarto Int. 1–21. doi: 10.1080/10106049.2019.1704070
Aouini, A., Matsukura, C., Ezura, H., and Asamizu, E. (2012). Characterisation of 13 glutamate receptor-like genes encoded in the tomato genome by structure, phylogeny and expression profiles. Gene 493, 36–43. doi: 10.1016/j.gene.2011.11.037
Arimura, G.-I. (2020). Making sense of the way plants sense herbivores. Trends Plant Sci. 26, 288–298. doi: 10.1016/j.tplants.2020.11.001
Arnemann, J. A., Roxburgh, S., Walsh, T., Guedes, J., Gordon, K., Smagghe, G., et al. (2019). Multiple incursion pathways for Helicoverpa armigera in Brazil show its genetic diversity spreading in a connected world. Sci. Rep. 9:19380. doi: 10.1038/s41598-019-55919-9
Azevedo, A. D., Prisco, J. T., Enéas-Filho, J., Abreu, C. E. B., and Gomes-Filho, E. (2006). Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 56, 87–94. doi: 10.1016/j.envexpbot.2005.01.008
Azevedo, R. A., Alas, R. M., Smith, R. J., and Lea, P. J. (1998). Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation, in the leaves and roots of wild-type and a catalase-deficient mutant of barley. Physiol. Plant 104, 280–292. doi: 10.1034/j.1399-3054.1998.1040217.x
Bar-On, Y. M., Phillips, R., and Milo, R. (2018). The biomass distribution on Earth. Proc. Natl. Acad. Sci. U.S.A. 115, 6506–6511. doi: 10.1073/pnas.1711842115
Bi, J. L., and Felton, G. W. (1995). Foliar oxidative stress and insect herbivory: Primary compounds, secondary metabolites, and reactive oxygen species as components of induced resistance. J. Chem. Ecol. 21, 1511–1530. doi: 10.1007/BF02035149
Bradford, M. M. (1976). Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3
Breeze, E. (2019). State of (In)flux: action of CNGC Ca2+ channel in defense against herbivory. Plant Cell 31, 1423–1424. doi: 10.1105/tpc.19.00372
Breiman, L., Friedman, J., Stone, C. J., and Olshen, R. A. (1984). Classification and Regression Trees. Boca Raton, FL: Chapman and Hall/CRC.
Burkepile, D. E., and Parker, J. D. (2017). Recent advances in plant-herbivore interactions. F1000Research 6:119. doi: 10.12688/f1000research.10313.1
Castro-Mercado, E., Martinez-Diaz, Y., Roman-Tehandon, N., and Garcia-Pineda, E. (2009). Biochemical analysis of reactive oxygen species production and antioxidative responses in unripe avocado (persea americana mill var hass) fruits in response to wounding. Protoplasma 235, 67–76. doi: 10.1007/s00709-009-0034-y
Chang, Y. W., Hsieh, C. J., Chang, K. W., Ringgaard, M., and Lin, C. J. (2010). Training and testing low-degree polynomial data mappings via linear SVM. J. Mach. Learn. Res. 11, 1471–1490. Available online at: https://www.jmlr.org/papers/volume11/chang10a/chang10a.pdf
Choi, W. G., Hilleary, R., Swanson, S. J., and Gilroy, S. (2016). Rapid, long-distance electrical and calcium signaling in plants. Annu. Rev. Plant Biol. 67, 287–307. doi: 10.1146/annurev-arplant-043015-112130
Choi, W. G., Miller, G., Wallace, I., Harper, J., Mittler, R., and Gilroy, S. (2017). Orchestrating rapid long-distance signaling in plants with Ca2+, ROS and electrical signals. Plant J. 90, 698–707. doi: 10.1111/tpj.13492
Cover, T., and Hart, P. (1967). Nearest neighbor pattern classification. IEEE Trans. Inf. Theory 13, 21–27. doi: 10.1109/TIT.1967.1053964
Cuin, T. A., Dreyer, I., and Michard, E. (2018). The role of potassium channels in arabidopsis thaliana long distance electrical signalling: AKT2 modulates tissue excitability while GORK shapes action potentials. Int. J. Mol. Sci. 19:926. doi: 10.3390/ijms19040926
Czepak, C., Albernaz, K. C., Vivan, L. M., Guimarães, H. O., and Carvalhais, T. (2013). Primeiro registro de ocorrência de Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) no Brasil. Pesqui. Agropecu. Trop. 43, 110–113. doi: 10.1590/S1983-40632013000100015
De Loof, A. (2016). The cell's self-generated “electrome”: the biophysical essence of the immaterial dimension of Life? Commun. Integr. Biol. 9;e1197446. doi: 10.1080/19420889.2016.1197446
de Medeiros, A. D., Capobiango, N. P., da Silva, J. M., da Silva, L. J., da Silva, C. B., and dos Santos Dias, D. C. F. (2020). Interactive machine learning for soybean seed and seedling quality classification. Sci. Rep. 10:11267. doi: 10.1038/s41598-020-68273-y
Duffey, S. S., and Stout, M. J. (1996). Antinutritive and toxic components of plant defense against insects. Arch. Insect Biochem. Physiol. 32, 3–37. doi: 10.1002/(SICI)1520-6327(1996)32:1<3::AID-ARCH2>3.0.CO2-1
Elstner, E. F., and Heupel, A. (1976). Inhibition of nitrite formation from hydroxylammonium-chloride: a simple assay for superoxide dismutase. Anal. Biochem. 70, 616–620. doi: 10.1016/0003-2697(76)90488-7
Ewing, N. N., and Bennett, A. B. (1994). Assessment of the number and expression of P-Type H+-ATPase genes in tomato. Plant Physiol. 106, 547–557. doi: 10.1104/pp.106.2.547
Fan, R. E., Chang, K. W., Hsieh, C. J., Wang, X. R., and Lin, C. J. (2008). LIBLINEAR: a library for large linear classification. J. Mach. Learn. Res. 9, 1871–1874. doi: 10.1145/1390681.1442794
Farmer, E. E., Gao, Y-Q., Lenzoni, G., Wolfender, J-L., and Wu, Q. (2020). Wound- and mechanostimulated electrical signals control hormone responses. New Phytol. 227, 1037–1050. doi: 10.1111/nph.16646
Fichman, Y., and Mittler, R. (2020). Rapid systemic signaling during abiotic and biotic stresses: is the ROS wave master of all trades? Plant J. 102, 887–896. doi: 10.1111/tpj.14685
Fitt, G. P. (1989). Ecology of Heliothis species in relation to agroecosystems. Annu. Rev. Entomol. 34, 17–52. doi: 10.1146/annurev.en.34.010189.000313
Forman, G., and Scholz, M. (2010). Apples-to-apples in cross-validation studies. SIGKDD Explor. 12, 49–57. doi: 10.1145/1882471.1882479
Freund, Y., and Schapire, R. E. (1997). A decision-theoretic generalization of on-line learning and an application to boosting. J. Comput. Syst. Sci. 55, 119–139. doi: 10.1006/jcss.1997.1504
Fromm, J., Hajirezaei, M.-R., Becker, V. K., and Lautner, S. (2013). Electrical signaling along the phloem and its physiological responses in the maize leaf. Front. Plant Sci. 4:239. doi: 10.3389/fpls.2013.00239
Gao, J., Zhang, Y., Li, Z., and Liu, M. (2020). Role of ethylene response factors (ERFs) in fruit ripening. FQS 1, 15–20. doi: 10.1093/fqsafe/fyz042
Giannopolitis, C. N., and Ries, S. K. (1977). Superoxide dismutases. I. Occurrence in higher plants. Plant Physiol. 59, 309–314. doi: 10.1104/pp.59.2.309
Hedrich, R., Salvador-Recatalà, V., and Dreyer, I. (2016). Electrical wiring and long-distance plant communication. Trends Plant Sci. 21, 376–387. doi: 10.1016/j.tplants.2016.01.016
Hoagland, D. R., and Arnon, D. I. (1938). The water culture method for growing plants without soil. Calif. Agric. Exp. Stn. 347, 1–39.
Hu, C., Wei, C., Ma, Q., Dong, H., Shi, K., Zhou, Y., et al. (2021). Ethylene response Factor 15 and 16 trigger jasmonate biosynthesis in tomato during herbivore resistance. Plant Physiol. 185, 1182–1197. doi: 10.1093/plphys/kiaa089
Huber, A. E., and Bauerle, T. L. (2016). Long-distance plant signaling pathways in response to multiple stressors: the gap in knowledge. J. Exp. Bot. 67, 2063–2079. doi: 10.1093/jxb/erw099
Kim, K. S., Choi, H. H., Moon, C. S., and Mun, C. W. (2011). Comparison of k-nearest neighbor, quadratic discriminant and linear discriminant analysis in classification of electromyogram signals based on the wrist-motion directions. Curr. Appl. Phys. 11, 740–745. doi: 10.1016/j.cap.2010.11.051
Kritikos, D. J., Ota, N., Hutchison, W. D., Beddow, J., Walsh, T., Tay, W. T., et al. (2015). The potential distribution of invading helicoverpa armigera in North America: is it just a matter of time? PLoS ONE 10:e0133224. doi: 10.1371/journal.pone.0133224
Kumari, A., Chételat, A., Nguyen, C. T., and Farmer, E. E. (2019). Arabidopsis H+-ATPaseAHA1 controls slow wave potential duration and wound-response jasmonate pathway activation. Proc. Natl. Acad. Sci. U.S.A. 116, 20226–20231. doi: 10.1073/pnas.1907379116
Liu, Y., and He, C. (2016). Regulation of plant reactive oxygen species (ROS) in stress responses: learning from AtRBOHD. Plant Cell Rep. 35, 995–1007. doi: 10.1007/s00299-016-1950-x
Morsomme, P., and Boutry, M. (2000). The plant plasma membrane H+-ATPase: structure, function and regulation. BBA Biomembr. 1465, 1–16. doi: 10.1016/S0005-2736(00)00128-0
Murúa, M. G., Scalora, F. S., Navarro, F. R., Cazado, L. E., Casmuz, A., Villagrán, M. E., et al. (2014). First record of Helicoverpa armigera (Lepidoptera: Noctuidae) in Argentina. Fla Entomol. 97, 854–856. doi: 10.1653/024.097.0279
Nakano, Y., and Asada, A. K. (1981). Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 22, 867–880.
Parise, A. G., Reissig, G. N., Basso, L. F., Senko, L. G. S., Oliveira, T. F. C., de Toledo, G. R. A., et al. (2021). Detection of different hosts from a distance alters the behaviour and bioelectrical activity of Cuscuta racemosa. Front. Plant Sci. 12:594195. doi: 10.3389/fpls.2021.594195
Pedreschi, R., and Lurie, S. (2015). Advances and current challenges in understanding postharvest abiotic stresses in perishables. Postharvest Biol. Technol. 107, 77–89. doi: 10.1016/j.postharvbio.2015.05.004
Pereira, D. R., Papa, J. P., Saraiva, G. F. R., and Souza, G. M. (2018). Automatic classification of plant electrophysiological responses to environmental stimuli using machine learning and interval arithmetic. Comput. Electron Agric. 145, 35–42. doi: 10.1016/j.compag.2017.12.024
Pratissoli, D., Lima, V. L. S., Pirovani, V. D., et al. (2015). Ocorrência de Helicoverpa armigera (Lepidoptera: Noctuidae) em tomateiro no estado do espírito Santo. Hortic. Bras. 33, 101–105. doi: 10.1590/S0102-053620150000100016
Rancic, D., and Pecinar, I. M. (2010). “Anatomy of tomato fruit and fruit pedicel during fruit development,” in Microscopy: Science, Technology, Applications and Education, eds A. Méndez-Vilas, J. Díaz (Badajoz: Formatex), 851–861.
Rasmussen, C. E., and Williams, C. K. I. (2006). Gaussian Processes for Machine Learning. Cambridge: The MIT Press. doi: 10.7551/mitpress/3206.001.0001
Reissig, G. N., Posso, D. A., Borella, J., da Silveira, R. V. D., Rombaldi, C. V., and Bacarin, M. A. (2018). High MT-sHSP23.6 expression increases antioxidant system in ‘Micro-Tom’ tomato fruits during post-harvest hypoxia. Sci. Hortic. 242, 127–136. doi: 10.1016/j.scienta.2018.07.035
Russell, S., and Norvig, P. (2003). Artificial Intelligence: A Modern Approach, 2nd Edn. Hoboken, NJ: Prentice Hall.
Saraiva, G. F. R., Ferreira, A. S., and Souza, G. M. (2017). Osmotic stress decreases complexity underlying the electrophysiological dynamic in soybean. Plant Biol. 19, 702–708. doi: 10.1111/plb.12576
Simmi, F.Z., Dallagnol, L.J., Ferreira, A.S., Pereira, D.R., and Souza, G.M. (2020). Electrome alterations in a plant-pathogen system: toward early diagnosis. Bioelectrochem 133:107493. doi: 10.1016/j.bioelechem.2020.107493
Souza, G. M., Ferreira, A. S., Saraiva, G. R., and Toledo, G. R. (2017). Plant “electrome” can be pushed toward a self-organized critical state by external cues: evidences from a study with soybean seedlings subject to diffrent environmental conditions. Plant Signal Behav. 12:e1290040. doi: 10.1080/15592324.2017.1290040
Souza, G. M., Toledo, G. R. A., and Saraiva, G. F. R. (2018). “Towards systemic view for plant learning: ecophysiological perspective,” in Memory and Learning in Plants. Signaling and Communication in Plants, eds F. Baluska, M. Gagliano, and G. Witzany (Springer), 163–189. doi: 10.1007/978-3-319-75596-0_9
Toledo, G. R. A., Parise, A. G., Simmi, F. Z., Costa, A. V. L., Senko, L. G. S., Debono, M.-W., et al. (2019). Plant electrome: the electrical dimension of plant life. Theor. Exp. Plant Physiol. 31, 21–46. doi: 10.1007/s40626-019-00145-x
Toyota, M., Spencer, D., Sawai-Toyota, S., Jiaqi, W., Zhang, T., Koo, A. J., et al. (2018). Glutamate triggers long-distance, calcium-based plant defense signaling. Science 361, 1112–1115. doi: 10.1126/science.aat7744
Tran, D., Dutoit, F., Najdenovska, E., Wallbridge, N., Plummer, C., Mazza, M., et al. (2019). Electrophysiological assessment of plant status outside a faraday cage using supervised machine learning. Sci. Rep. 9:17073. doi: 10.1038/s41598-019-53675-4
Valoy, M., López-Acosta, J. C., Lomáscolo, S., Bernacki,1, F., Varela, O., and Ordano, M. (2019). Leaf herbivory modulates fruit trait correlations within individual plants. Arthropod Plant Inte. 14, 373–385. doi: 10.1007/s11829-020-09740-1
Velikova, V., Yordanov, I., and Edreva, A. (2000). Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Protective role of exogenous polyamines. Plant Sci. 151, 59–66. doi: 10.1016/S0168-9452(99)00197-1
Vilela, M., Mendes, S. M., Valicente, F. H., Carvalho, S. D., dos Santos, A. E., dos Santos, C. A., et al. (2014). Metodologia para criação e manutenção de Helicoverpa armigera em laboratório. Circular Técnica. Sete Lagoas: Embrapa Milho e Sorgo.
Vodeneev, V. A., Katicheva, L. A., and Sukhov, V. S. (2016). Electrical signals in higher plants: mechanisms of generation and propagation. Biophysics 61, 505–512. doi: 10.1134/S0006350916030209
War, A. R., Paulraj, M. G., Ahmad, T., Buhroo, A. A., Hussain, B., Ignacimuthu, S., et al. (2012). Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 7, 1306–1320. doi: 10.4161/psb.21663
Windeatt, T. (2006). Accuracy/diversity and ensemble MLP classifier design. IEEE Trans Neur. Netw. 17, 1194–1211. doi: 10.1109/TNN.2006.875979
Zhou, H., Bai, S., Wang, N., Sun, X., Zhang, Y., Zhu, J., et al. (2020). CRISPR/Cas9-mediated mutagenesis of MdCNGC2 in apple callus and VIGS-mediated silencing of MdCNGC2 in fruits improve resistance to Botryosphaeria dothidea. Front. Plant Sci. 11:575477. doi: 10.3389/fpls.2020.575477
Keywords: plant electrophysiology, machine learning, Micro-Tom, Helicoverpa amigera, stress response, antioxidant system
Citation: Reissig GN, Oliveira TFC, Oliveira RP, Posso DA, Parise AG, Nava DE and Souza GM (2021) Fruit Herbivory Alters Plant Electrome: Evidence for Fruit-Shoot Long-Distance Electrical Signaling in Tomato Plants. Front. Sustain. Food Syst. 5:657401. doi: 10.3389/fsufs.2021.657401
Received: 22 January 2021; Accepted: 21 June 2021;
Published: 20 July 2021.
Edited by:
Daniel Tran, Institute for Plant Production Systems–Agroscope, SwitzerlandReviewed by:
Ken Yokawa, Kitami Institute of Technology, JapanElena Najdenovska, University of Applied Sciences and Arts of Western Switzerland (HES-SO), Switzerland
Copyright © 2021 Reissig, Oliveira, Oliveira, Posso, Parise, Nava and Souza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gabriela Niemeyer Reissig, gabriela.niemeyer.reissig@gmail.com